Le système immunitaire n’est jamais
“en pause” — épisode 2
Si tu as déjà entendu que le système immunitaire est
parfois “en pause” ou “en sommeil” et que cet état serait un problème car il
aurait “du mal à se réveiller” et que cela nous exposerait à des maladies plus
fréquentes et plus délétères, alors ce billet est fait pour toi. On va voir
ensemble pour quelles raisons cette assertion relève d’un “bon sens” naïf, qui
ne tient plus la route une fois qu’on en sait plus sur les mécanismes et des
processus immunitaires et leur diversité. On y va ? Prépare-toi, le voyage
sera long ! Et cet épisode-ci est effectivement assez long.
L’épisode
1, c’est par là.
Confiné
conceptuellement à son seul rôle d’élimination d’organismes pathogènes, on
pourrait faussement croire que le système immunitaire ne sert
qu’occasionnellement, c’est-à-dire seulement lorsque nous sommes malades,
envahis par un microorganisme néfaste, et que le reste du temps, il est “en
pause” ou “en sommeil”. C’est faux.
Le système immunitaire fonctionne en permanence et n’est jamais “en sommeil”[1],
comme on l’a déjà vu au travers de ses multiples rôles physiologiques
permanents et variés. La croyance infondée qu’il aurait de ce fait besoin
d’être “stimulé” (boosté) soit par
des infections, soit par des compléments est ainsi assez répandue, surtout dans
le second cas par des pages Internet à visée commerciale[2].
Tu
pourrais contre-argumenter, cher lecteur, chère lectrice, que la détection de
cellules mortes, endommagées et sénescentes ou la régulation de l’homéostasie
sont des choses normales, de base, et
qu’on pourrait considérer que l’arrivée d’une entité étrangère, comme un
microorganisme pathogène, déclenche d’autres mécanismes spécifiques qui, eux,
seraient “en sommeil” lorsqu’ils sont absents. Mais tu aurais tort. Pour
comprendre pour quelle raison, il va falloir qu’on parle de la manière dont le
système immunitaire détecte les entités étrangères dans nos organismes, la
manière dont il différencie le “soi” (qui fait partie de l’organisme) de ce qui
est étranger (le “non soi”) ou anormal. Tu verras que la détection d’organismes
pathogènes se produit de la même façon que pour les microorganismes résidents
permanents (le microbiote) ou que pour les cellules du soi anormales, et
qu’elle déclenche les mêmes processus
immunitaires.
 |
| Figure 1 |
Il n’y a pas de “sous-stimulation”
du système immunitaire en raison d’un “manque d’exposition aux pathogènes”,
car cela sous-entendrait qu’il existe des mécanismes de reconnaissance
différents pour deux catégories concrètes d’organismes : les pathogènes et
les non-pathogènes — catégories sémantiques qui n’ont pas d’existence réelle,
puisque la pathogénie — on en a déjà parlé ici — est contextuelle et que ce qui est reconnu
par le système immunitaire ne diffère pas entre pathogènes et non-pathogènes.
Encore une fois, ce qui est reconnu, ce sont des caractéristiques anormales. Il existe des millions d’espèces de microorganismes[3],
et très peu sont pathogènes. Nous les croisons en permanence, partout, et il est
impossible de les éviter: ils sont dans l’air, dans l’eau, dans les aliments,
sur la peau — partout. Pourquoi ne nous envahissent-ils pas, alors que nous
représentons pour eux une source gigantesque de nutriments et un havre
potentiel de vie calme et tranquille (Fig. 1), loin des affres changeants du
monde extérieur?[4]
Pourquoi ne sont-ils pas pathogènes ?
Barrières physiques et chimiques
Il
y a plusieurs raisons[5][6]:
premièrement, les conditions internes de nos corps ne conviennent pas à leur
survie ou à leur développement (l’humidité, la composition de nos fluides
biologiques, la salinité, le pH, la température, la présence ou non d’oxygène,
etc.), et on ne les y trouve jamais. Ils ne peuvent donc pas proliférer (Fig. 2)
ni, de ce fait, provoquer d’effets néfastes. C’est déjà une première barrière
sélective qui contribue à l’immunité dont nous bénéficions face à eux. Elle ne
nécessite ni détection particulière, ni cellules spécialisées. C’est simplement
une immunité de base conférée par la simple pression de sélection exercée sur
les microorganismes par nos propres conditions physiologiques internes. On en a
déjà parlé par là.
 |
Figure 2: La croissance microbienne, l’exemple d’une bactérie. Ici, l’exemple est celui d’un bacille comme Escherichia coli ou Salmonella
enterica. Attention, dans le cas des virus, qui ne sont pas autonomes, ceci
n’est pas valable. |
Deuxièmement,
certains microorganismes sont incapables de franchir les barrières physiques
(peau, muqueuses, etc.) qui leur permettraient d’entrer à l’intérieur de nos
corps. Le pH acide de l’estomac est, par exemple, une condition
physico-chimique qui exerce une pression de sélection considérable sur les
microorganismes qui transitent par notre tube digestif depuis la bouche. Très
peu de microorganismes sont capables d’y survivre et encore moins de s’y
développer[7].
La peau, les muqueuses, toutes ces barrières constituent également des obstacles
physiques que peu de microorganismes parviennent à franchir.
Troisièmement,
le système immunitaire — tel qu’on
l’entend habituellement — existe. Et il
n’est pas passif en attendant que des
microorganismes entrent, comme s’ils étaient rares et n’apparaissaient
qu’occasionnellement. Non. Ils sont présents partout, tout le temps.
Le système immunitaire inné est notre “première ligne de défense” s’il on veut
utiliser un champ lexical militaire. En plus de son intervention dans pléthore
de mécanismes physiologiques (voir par là), ce dernier est aussi
capable d’empêcher la prolifération des microorganismes. Si les conditions
physiologiques de nos corps sont déjà une barrière sélective empêchant la
survie et/ou la prolifération de très nombreux microorganismes, le système
immunitaire est aussi capable de les modifier et les rendre impropres à la
survie d’une partie des microorganismes étrangers qui les toléreraient malgré
tout. Autrement dit, le système immunitaire agit comme une “machine à pressions
de sélection” vis-à-vis des organismes étrangers (c’est ce que je te présentais ici). Ces
conditions délétères pour les visiteurs inopportuns le sont aussi pour nos
propres cellules et les tissus qu’elles constituent, et ne persistent que pour
une durée normalement limitée.
Si
l’augmentation de la température corporelle (fièvre[8])
limite parfois la croissance microbienne[9][10]
(et de loin pas dans tous les cas), ce n’est pas une barrière physique
suffisante[11], mais surtout une manifestation de
l’augmentation du métabolisme et de la thermogenèse — potentialisation du
métabolisme indispensable aux cellules immunitaires spécialisées en pleine
expansion pendant une infection. Dit autrement, la machinerie doit tourner à
plein. Il y a aussi — et c’est souvent par là que commencent les défenses — ce
qu’on appelle l’ “immunité nutritionnelle” au cours de laquelle les
cellules immunitaires comme les neutrophiles ou les macrophages sécrètent des
protéines qui piègent et séquestrent tous les métaux essentiels tels que fer, zinc,
cuivre, etc[12][13][14]
qui ne sont dès lors plus disponibles non plus pour les microorganismes qui en
ont autant besoin que nous (on en a parlé là).
Ces
barrières physiques et chimiques, ainsi que ces mécanismes de limitation des
nutriments permettent d’appliquer une grande série de contraintes
physico-chimiques et biologiques qui changent le paradis à microorganismes que représente notre corps en une sorte de désert
aride, où les ressources sont inaccessibles et les conditions de survie
défavorables. Cela peut empêcher la pullulation de ces microorganismes. Mais si
elles sont efficaces contre la très grande majorité des micro-organismes que
nous croisons, ces conditions variables ne suffisent manifestement pas
toujours.
Systèmes
immunitaires inné et adaptatif
Conventionnellement, on a coutume de séparer le
système immunitaire en deux sous-ensembles[15] :
le système immunitaire inné et
le système immunitaire adaptatif.
Le premier est présent chez tous les animaux (et il en existe aussi des
équivalents chez les plantes, les champignons, les eucaryotes unicellulaires,
ainsi que chez les bactéries), tandis que le second n’est apparu que chez les
vertébrés dont nous faisons partie[16][17][18][19]. Ces
deux “systèmes” ne sont pas ontologiquement distincts, c’est-à-dire qu’ils ne
sont pas deux entités bien délimitées et existant indépendamment l’une de
l’autre, qu’on pourrait séparer ; le second ne va pas sans le premier. En
fait, comme le système immunitaire adaptatif est apparu après le système immunitaire inné (il y a environ 500 millions
d’années[20]),
son fonctionnement repose sur les fondements du plus ancien et est intimement
intriqué avec lui. Certes, le système immunitaire adaptatif dispose
d’innovations absentes chez l’autre, notamment dans la manière dont il détecte
les pathogènes et les entités n’appartenant pas au soi, et dans sa façon
d’acquérir de nouvelles capacités de reconnaissance spécifiques (sa mémoire),
mais il s’est construit avec le
système immunitaire inné, et pas indépendamment de lui. En d’autres termes, ils
ont co-évolué[21][22][23].
La frontière entre les deux n’existe donc pas réellement, elle est
conceptuelle. Il faut donc se garder de penser qu’ils sont indépendants.
Je ne parlerai pour l’instant que de système
immunitaire inné, surtout parce que c’est lui qui intervient toujours en
premier, rapidement, lorsqu’une entité étrangère parvient à entrer dans notre
organisme[24].
C’est d’ailleurs aussi le seul à pouvoir fonctionner immédiatement lorsqu’on
rencontre un microorganisme pour la première fois et que l’immunité adaptative
ne permet pas encore de le reconnaître spécifiquement. C’est le système
immunitaire inné qui intervient dans la reconnaissance immédiate des organismes
étrangers, mais aussi dans la détection des cellules anormales, mortes ou
sénescentes, etc. Il est aussi actif en permanence pour maîtriser le
microbiote.
C’est donc de lui qu’il faut parler en premier.
Des cellules
spécialisées
Le système immunitaire
est le nom qu’on donne à tout un ensemble de processus, d’organes et de
cellules très différents. Souviens-toi que cette machinerie complexe est aussi
impliquée dans beaucoup de processus physiologiques qui ne sont pas directement
reliés à la “défense contre les pathogènes”. Mais, en général, lorsqu’on parle
du système immunitaire, on fait référence à sa fonction de défense et, surtout,
aux cellules spécialisées qui en sont
les acteurs. Le système immunitaire inné, par exemple, possède son ensemble de
cellules spécialisées, lesquelles sont en général unicellulaires (elles circulent librement !)[25]:
les macrophages, les microglies qui sont des formes de
macrophages du système nerveux central, les mastocytes, les polynucléaires
neutrophiles, basophiles ou éosinophiles, les cellules dendritiques, les monocytes,
les cellules lymphoïdes innées[26][27]
(ILC, pour Innate lymphoid cells)
telles que les ILC-1, ILC-2, ILC-3, les cellules Natural Killer (NK), les LTi), les lymphocytes T invariants associés aux muqueuses (dits MAIT, Mucosal-associated invariant T cells) et
les lymphocytes T gd (gamma-delta)... et beaucoup d'autres. Du monde, donc (Figure 3).
Le système immunitaire
adaptatif lui aussi possède des cellules spécialisées, telles que les lymphocytes B (producteurs
d’anticorps), les plasmocytes (forme
différentiée de lymphocytes B produisant les anticorps circulants), et la
famille des lymphocytes T (dits ab),
comme les lymphocytes T cytotoxiques
(TC), les lymphocytes T Natural Killer (TNK), les lymphocytes T auxiliaires, dits helper
(Th), avec les sous-types Th1, Th2, Th17, Th9, Th22, Tfh), les lymphocytes T régulateurs (Treg),
mais on peut aussi compter les lymphocytes
B mémoires et les lymphocytes T
mémoires (plusieurs sous-types : les TCM, TEM, TRM,
TVM), qui donnent corps à la fameuse “mémoire adaptative” (Figure 3).
 |
Figure 3: Les organes et les cellules immunitaires spécialisés. La figure présente les organes du système immunitaire et qui participent directement à ce qu'on appelle "la défense contre les pathogènes" (production et maturation des cellules immunitaires, etc.), mais aussi les barrières qui participent à la maîtrise des entrées d'entités étrangères (peau, muqueuses, etc.). Enfin, la partie droite présente des exemples de cellules immunitaires spécialisées, comme certaines de celles appartenant au système immunitaire dit "inné" (cellules natural killer, polynucléaires, mastocytes, lymphocytes T invariants, lymphocytes T gamma delta, cellules lymphoïdes, etc.), et certaines appartenant au système immunitaire dit "adaptatif" (lymphocytes B et T "alpha beta", cytotoxiques, auxiliaires, régulateurs, natural killer, plasmocytes, etc.).
|
Toutes ces cellules
naissent dans la moelle osseuse et voyagent dans différents organes où elles
suivent leur trajectoire de maturation et où elles assurent ensuite beaucoup de
fonctions: rate, thymus, nœuds (ganglions) lymphatiques, vaisseaux
lymphatiques, amygdales, tissus lymphoïdes, etc. sont tous des organes qu’on associe
directement au système immunitaire. Ces cellules immunitaires se distribuent
aussi dans de nombreux autres endroits : sang, organes, tissus lymphoïdes,
etc. Il en existe plusieurs centaines de milliards à tout moment chez un
individu humain moyen, représentant près d’1,2 kg en termes de masse corporelle[28].
Avec les cellules sanguines telles que les hématies (globules rouges) ou les
plaquettes dérivées de cellules sanguines, elles sont parmi les types
cellulaires les plus abondants[29].
Mais attention, ne va pas croire que ces
cellules spécialisées sont les seules à contribuer au système immunitaire.
L’immunité innée, par exemple, fait intervenir d’autres cellules qui ne sont
pas à proprement parler des cellules immunitaires. Les cellules épithéliales
(celles qui constituent les tissus, et qui sont associées entre elles pour
former des surfaces) et les cellules endothéliales (celles qui forment les
vaisseaux sanguins, par exemple) sont directement impliquées dans l’immunité
innée. Par exemple, les cellules épithéliales intestinales, qui forment les
surfaces des intestins, participent à la détection des microorganismes qui
passent par le tube digestif, et répondent à leur présence. De manière
générale, toutes les cellules des surfaces mucosales (des muqueuses),
c’est-à-dire en contact avec le milieu extérieur (bouche, tube digestif, donc
intestins, rectum, vagin, voies urinaires, voies respiratoires, etc.),
contribuent au système immunitaire inné, et, indirectement, au système
immunitaire adaptatif[30].
Un concept important: les récepteurs
Lorsqu’on
dit que “le système immunitaire perçoit”
la présence d’anomalies, ce qu’un biologiste dit en réalité, c’est que ce sont
entre autres toutes ces cellules qui, d’une manière ou d’une autre, “détectent”
les corps étrangers, les microorganismes et/ou les anomalies. On va donc
maintenant voir comment cela se produit.

Les
cellules du système immunitaire ne reconnaissent pas spécifiquement les
“organismes pathogènes” — c’est une idée fausse[31]
— car, encore une fois, il n’y a pas de caractéristiques propres aux
“pathogènes”[32];
elles détectent tout ce qui n’est pas le
soi, autrement dit aussi bien des microorganismes dangereux et pathogènes
que les autres, incluant de simples molécules, telles que les toxines. Cette
reconnaissance est possible parce que nos cellules disposent de “détecteurs”,
des récepteurs en langage
plus formel[33],
capables d’interagir (former une association à l’échelle moléculaire) avec des
structures qui ne font pas partie du
“soi”, permettant ensuite aux cellules d’y réagir par une réponse appropriée.
C’est ici, donc, qu’il va nous falloir discuter de la notion de récepteur,
fondamentale en biologie.
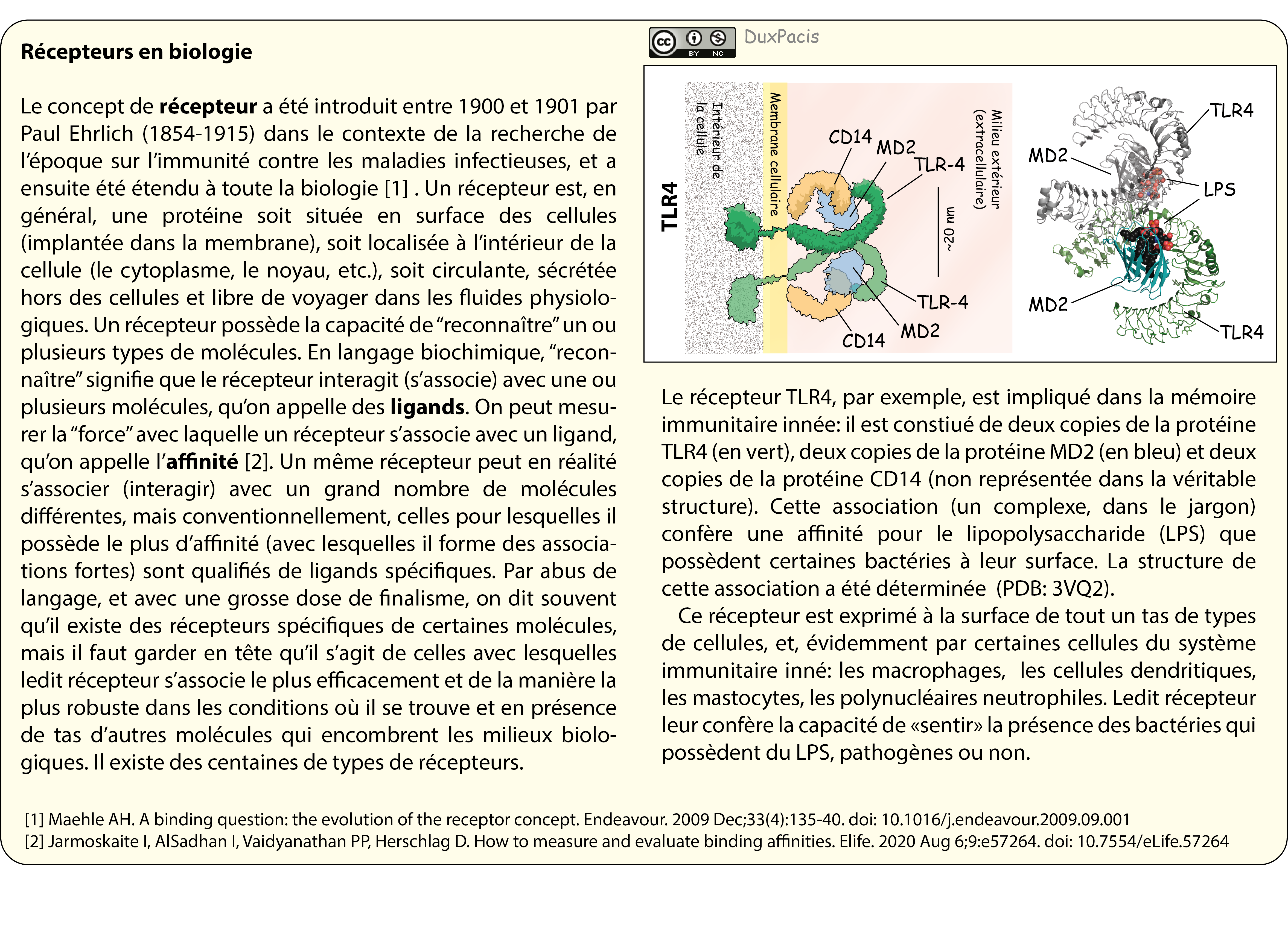
Le
concept de récepteur a été introduit entre 1900 et 1901 par Paul Ehrlich
(1854-1915)[34],
et a ensuite été étendu à toute la biologie. Un récepteur est, en général, une protéine soit située en surface
des cellules (implantée dans la membrane), soit localisée à l’intérieur de la
cellule (le cytoplasme, le noyau, etc.), soit circulante, sécrétée hors des
cellules et libre de voyager dans les fluides physiologiques. Un récepteur
possède la capacité de “reconnaître” un ou plusieurs types de molécules. En
langage biochimique, “reconnaître” signifie que le récepteur interagit avec une ou plusieurs molécules,
qu’on appelle des ligands.
On peut mesurer la “force” avec laquelle un récepteur s’associe avec un ligand,
qu’on appelle l’affinité[35].
Un même récepteur peut en réalité s’associer (interagir) avec un grand nombre
de molécules différentes, mais conventionnellement,
celles pour lesquelles il possède le plus d’affinité (avec lesquelles il forme
des associations fortes) sont appelées ligands. Par abus de langage, on dit
souvent qu’il existe des récepteurs spécifiques
de certaines molécules, mais il faut garder en tête qu’il s’agit de celles avec
lesquelles il s’associe le plus efficacement et de la manière la plus robuste dans les conditions où il se trouve et
en présence de tas d’autres molécules qui encombrent les milieux biologiques.
Il
existe des milliers de récepteurs différents. Dans le cas du système
immunitaire inné, on les appelle récepteurs
innés. Ils interagissent avec des molécules le plus souvent présentes à la
surface des microorganismes ou des cellules dites anormales, c’est-à-dire des
structures qui n’existent normalement pas chez nous.
L’association
entre un récepteur et un ligand est la première étape d’une détection, mais elle n’est pas
suffisante, car il faut ensuite que cette association déclenche des mécanismes
de réponse, une cascade d’évènements moléculaires qui modifient l’état
physiologique de la cellule possédant ce récepteur (généralement en changeant
la manière dont les gènes sont exprimés). C’est ce qu’on appelle une voie de transduction du signal.
J’appellerai cela une réponse, pour
simplifier, mais retiens qu’il s’agit d’un réseau complexe d’évènements
moléculaires en cascade qui non seulement signent la détection d’un ligand par
le récepteur, mais aussi l’amplifie. Il existe beaucoup de voies de
transduction du signal différentes selon les récepteurs impliqués, le type de
cellule, etc. De même, chaque type de cellule humaine exprime des combinaisons
de récepteurs qui lui sont propres — c’est d’ailleurs l’un des moyens de
distinguer les différents types de cellules entre elles.
Le
système immunitaire est aussi impliqué dans la reconnaissance des cellules du
soi “anormales” et perçoit, selon le même principe et avec des récepteurs
dédiés, des caractéristiques normalement absentes (ou masquées)[36][37].
Si tu veux, les récepteurs sont des “organes sensitifs” cellulaires.
 Nous disposons de
récepteurs aux molécules qui permettent de sentir ce que nous appelons des
odeurs[38]
ou des goûts[39],
des récepteurs aux sons[40],
des récepteurs à la lumière[41],
même des récepteurs à la chaleur[42]
ou à la pression[43][44]
et qui nous confèrent nos sens (odorat, goût, ouïe, vue, toucher, mais aussi
proprioception, thermoception, nociception, etc.). Nous sommes équipés
également de récepteurs à caractéristiques moléculaires anormales.
Nous disposons de
récepteurs aux molécules qui permettent de sentir ce que nous appelons des
odeurs[38]
ou des goûts[39],
des récepteurs aux sons[40],
des récepteurs à la lumière[41],
même des récepteurs à la chaleur[42]
ou à la pression[43][44]
et qui nous confèrent nos sens (odorat, goût, ouïe, vue, toucher, mais aussi
proprioception, thermoception, nociception, etc.). Nous sommes équipés
également de récepteurs à caractéristiques moléculaires anormales.
Mais qu’est-ce que le système immunitaire détecte, au
juste ? Et comment ?
Les récepteurs de l’immunité innée
Nos
cellules sont équipées de tout un tas de récepteurs (Fig. 4), c’est-à-dire de protéines de surface, ancrées dans la
membrane cellulaire et exposées à l’extérieur, mais aussi de protéines
internes, présentes dans les
cellules, et qui peuvent interagir avec (reconnaître) des structures
caractéristiques et initier tout un mécanisme de signalisation cellulaire dont
l’aboutissement est le déclenchement d’une réponse
immunitaire — un phénomène de cascade d’évènements moléculaires qu’on appelle l’inflammation[45].
Note
que le terme inflammation n’a pas ici
le sens négatif qu’on lui confère dans le langage courant, où il est associé à
la douleur, le gonflement et rougeoiement dû à une blessure — sens négatif qui
a longtemps été la norme, même en immunologie, lorsqu’on pensait au XIXe siècle
qu’elle était due exclusivement aux pathogènes ou aux états pathologiques,
alors que les découvertes ultérieures ont montré que ce phénomène physiologique
est déclenché, contrôlé et régulé par le système immunitaire lui-même[46].
Avec la découverte des mécanismes sous-jacents, il est devenu clair qu’ils
étaient communs à toutes les réponses immunitaires, d’où le fait
qu’aujourd’hui, inflammation se réfère en biologie et en médecine aux mécanismes de
mise en route de la réponse immunitaire. En fait, on connaît aujourd’hui
trois types différents d’inflammation, trois types de réponses immunitaires
selon les cellules impliquées dans la détection d’anomalies, les anomalies en
question, et la réponse qu’elles produisent[47].
 Les cellules du système
immunitaire inné, telles que les macrophages, les polynucléaires neutrophiles,
les cellules dendritiques, etc. sont bien entendu équipées de tels récepteurs,
mais ils sont aussi exprimés dans des cellules qui ne sont pas spécialisées
dans l’immunité, comme les cellules qu’on retrouve dans les parois délimitant
les vaisseaux (cellules endothéliales), les organes et les tissus (cellules
épithéliales, fibroblastes, etc.) (voir Fig. 4)[48].
Les cellules du système
immunitaire inné, telles que les macrophages, les polynucléaires neutrophiles,
les cellules dendritiques, etc. sont bien entendu équipées de tels récepteurs,
mais ils sont aussi exprimés dans des cellules qui ne sont pas spécialisées
dans l’immunité, comme les cellules qu’on retrouve dans les parois délimitant
les vaisseaux (cellules endothéliales), les organes et les tissus (cellules
épithéliales, fibroblastes, etc.) (voir Fig. 4)[48].
 |
| Figure 4: Que sont un épithélium, un endothélium, un récepteur ? Le panneau en haut à
gauche explique ce que sont les épithélia (un épithélium, des épithélia) et les
endothélia (un endothélium, des endothélia), le panneau en bas à gauche donne
l’exemple de l’épithélium qui constitue la surface de la paroi nasale. Les deux
panneaux de droite expliquent ce que sont les récepteurs, et la manière dont
ils « détectent » les anomalies (ici les molécules anormales), en
prenant l’exemple (en bas) du récepteur TLR4 qui reconnaît, entre autres
choses, le lipopolysaccharide caractéristique de toute une famille de
bactéries. |
Tu vois donc déjà ici que le “système immunitaire inné” ne comprend pas que les cellules immunitaires
spécialisées, mais toutes les cellules
capables de détecter des anomalies par l’entremise de ces récepteurs et de
signaler leur reconnaissance. C’est donc un système qui se confond avec presque
toutes les cellules les plus abondantes de notre organisme. Note aussi que
certains de ces récepteurs n’ont pas exclusivement un rôle immunitaire,
certains d’entre eux étant des acteurs centraux lors du développement
embryonnaire et la mise en place des organes[49].
Un exemple supplémentaire de fonction multitâche (on dit fonctions moonlighting, dans le jargon). C’est
aussi un nouveau témoignage, parmi des milliers d’exemples, que les systèmes
biologiques ne sont pas constitués de modules bien délimités mais des réseaux
qui s’entrecroisent, se confondent, interagissent. Et une illustration du fait
que les entités biologiques n’ont jamais qu’une seule fonction dédiée, mais
qu’elles peuvent en réalité participer à des tas de processus, sans hiérarchie
particulière.
Tous
les animaux — nous compris — disposent d’une collection de base de récepteurs innés codés sous la forme
de gènes et exprimés en protéines dédiées à la reconnaissance de structures
moléculaires. C’est une mémoire fixe,
qu’on peut qualifier de “prédéterminée”
et transmise génétiquement de génération en génération (d’où le “inné”). Elle
n’est pas modifiable et reste la même de la naissance à la mort[50][51].
Il existe, bien entendu, des différences entre
les individus et les populations, selon l’histoire des mutations survenues dans
les lignées humaines, y compris à l’échelle familiale — des polymorphismes,
dans le jargon — mais ils ne changent pas chez
un individu donné au cours de sa vie. Ces différences, en revanche, sont à
l’origine de variations dans la manière dont le système immunitaire inné répond
aux anomalies[52][53][54].
La
reconnaissance des entités étrangères ou anormales est un sujet complexe et
assez dense, et je ferai un billet dédié à cette thématique, mais je peux en
revanche te donner une idée de ce dont il retourne, et te montrer pour quelle
raison prétendre que seuls les organismes pathogènes “stimulent” le système
immunitaire est faux.
 |
Figure 5: Les caractéristiques moléculaires microbiennes. Les récepteurs de l’immunité innée
reconnaissent des caractéristiques conservées chez tous les microorganismes.
Ici, je donne l’exemple des bactéries. Il existe deux grandes familles de
bactéries, dont la première constitue les bactéries didermes (littéralement « avec deux peaux »), qui
possèdent deux membranes. Là, j’effectue une coupe imaginaire de l’une de ces
bactéries pour que tu puisses voir à l’intérieur. Elles possèdent une membrane
plasmique, comme toutes les cellules, constituée de lipides, mais aussi une
seconde membrane, la membrane externe. Elle est surtout composée d’un lipide
complexe, le lipide A, qui est décoré de nombreux sucres (des saccharides) dont
je n’ai représenté ici que deux exemples. En réalité, le lipide A peut être
agrémenté d’un grand nombre de ces molécules. L’ensemble lipide A + saccharides
est appelé le lipopolysaccharide (LPS). Il existe chez la majorité des
bactéries à deux membranes (aussi appelées à Gram-négatif, car elles ne se
colorent pas au test de Gram). Les bactéries Escherichia coli, Salmonella
enterica, Pseudomonas aeruginosa
et bien d’autres possèdent ce lipide. Le récepteur de l’immunité innée TLR4
reconnaît la partie lipide A. Il peut donc « détecter » toutes les
bactéries de ce type, qu’elles soient pathogènes ou non. L’autre type de
bactérie est représenté par les monodermes, qui ne possèdent qu’une seule
membrane, la membrane plasmique. Comme les didermes, elles possèdent une paroi,
le peptidoglycane, mais il est ici en général beaucoup plus épais et directement
exposé à l’extérieur (pas de membrane externe). Certains lipides de la membrane
plasmique sont décorés de sucres et d’acide lipoteichoïque, lequel est
« détecté » par les Toll-like Receptors (TLR) 2 et 4. Les bactéries
comme Staphylococcus aureus (Staphylocoque
doré), Streptococcus pneumoniae et
bien d’autres possèdent ce genre de structures, qu’ils soient pathogènes ou
non. Le peptidoglycane, quant à lui, peut être reconnu par des récepteurs
appelés NOD-like receptors. Il existe aussi des récepteurs capables de détecter
la présence de certaines protéines de surface des bactéries, telles les
flagellines (constituants des flagelles de certaines bactéries) reconnues par
le Toll-like receptor 5 (TLR5). |
Les récepteurs innés reconnaissent
des caractéristiques générales présentes chez les microorganismes.
Ils ne distinguent pas les espèces de microorganismes, ni s’ils sont pathogènes
ou non, mais détectent des constituants conservés
(c’est-à-dire qui sont partagés par beaucoup de microorganismes). On appelle cela
des Microbe-Associated
Molecular Patterns (MAMPs),
qu’on pourrait traduire par signatures
moléculaires associées aux microbes. Les récepteurs innés sont de ce fait
appelés des Pattern Recognition Receptors (PRR, récepteurs de
reconnaissance de patrons/signatures). Pour les bactéries, par exemple, nos
cellules disposent de plusieurs récepteurs reconnaissant certaines de leurs
molécules de surface (Fig. 5) :
-
Le peptidoglycane qui
joue chez les bactéries le rôle d’une charpente cellulaire[55].
Il est détecté par des récepteurs intracellulaires (à l’intérieur de nos
cellules, donc) appelées NOD-like receptors (NLR)[56][57][58].
Le peptidoglycane, bien qu’ayant des caractéristiques variables, possède une
structure de base conservée, et il est présent chez un très grand nombre de
bactéries (très rares sont celles qui en sont dépourvues). C’est cette
structure de base, ce squelette moléculaire commun, qui est reconnu par les
NLR.
- Le lipopolysaccharide[59][60]
est un lipide complexe de la membrane de certaines bactéries (dites à
Gram-négatif, voir Fig. 5). Il peut être très variable entre les
bactéries, mais il possède un constituant de base commun à toutes les bactéries
à Gram négatif : le lipide A (Fig. 5). Il est détecté par des
récepteurs exposés à la surface de nos cellules appelés Toll-like receptor 4
(TLR4)[61].
- Certains autres lipides complexes ou
protéines bactériennes de surface sont aussi des marqueurs de reconnaissance
par plusieurs récepteurs TLR différents (TLR1, 2, 5, et 6)[62].
La flagelline, par
exemple, est une protéine qui constitue les flagelles bactériens (Fig. 5),
qui permettent à ces cellules de se déplacer[63].
Elle est conservée chez de nombreuses bactéries et est reconnue par le
récepteur inné appelé TLR5[64].
- Pour les champignons, la détection passe
plutôt par des sucres complexes constitutifs de leurs parois cellulaires qui n’ont pas d’équivalent chez nous, qu’on
appelle des polysaccharides[65].
Il en existe plusieurs familles différentes[66], reconnues par plusieurs
récepteurs différents, comme les C-type lectin receptors (CLRs)[67].
- Dans le cas des virus, la reconnaissance
innée est généralement (mais pas exclusivement) intracellulaire, et se produit
lorsqu’il a déjà envahi une cellule cible. Comme un virus possède peu de
composants, et que ces derniers sont produits par ou “volés” à nos cellules, la
détection est beaucoup plus difficile, car, en termes de constituants, ils nous
ressemblent de fait. Souvent, la
reconnaissance d’une infection virale passe par la détection du génome viral
(ADN ou ARN) dont les caractéristiques et/ou la localisation à l’intérieur de
nos cellules suffisent à les différencier de nos propres constituants[68].
Certains récepteurs TLR (TLR3, 7, 8, 9, 10) peuvent reconnaître la présence de
protéines, d’ADN ou d’ARN viraux, mais d’autres récepteurs (RIG-I, MDA5, LGP2),
peuvent aussi s’acquitter de cette tâche à différents endroits de nos cellules
et à différents stades du cycle viral.
L’ensemble
de ce processus de détection d’entités étrangères par les récepteurs innés et
des mécanismes conduisant à son élimination ou à son atténuation est appelée réponse immunitaire innée[69].
La Fig.
6 présente quelques récepteurs de l’immunité innée.
 |
| Figure 6: Récepteurs de l’immunité innée. Le tableau donne
quelques exemples (la liste n’est pas du tout exhaustive !) de récepteurs
de l’immunité innée chez les humains. Ici, il s’agit des Toll-like récepteurs
(TLR). Chaque protéine TLR est codée par son propre gène. La protéine TLR1 est
codée par un gène, la protéine TLR2 par un autre, et ainsi de suite. Les
récepteurs TLR fonctionnels résultent de l’assemblage de deux protéines
TLR : soit deux fois la même (on parle alors d’un homodimère), comme dans
le cas de TLR3, 4 et 5), soit il s’agit de l’association de deux protéines TLR
différentes (un hétérodimère), comme pour TLR1 et TLR2 ou encore TLR2 et TLR6.
La protéine TLR2, par exemple, s’associe avec TLR1, et lorsque c’est le cas, le
récepteur qui en résulte reconnaît des triacyl-lipopeptides, des structures qui
existent à la surface de certaines bactéries. Lorsqu’elle s’associe à TLR6, le
complexe (hétérodimère) TLR2/TLR6 reconnaît alors tout une série de signatures
microbiennes bactériennes et fongiques (de champignons). Le tableau présente 5
récepteurs de type TLR, leur structure (les codes de la Protein Data Bank, PDB,
où ces structures sont stockées et accessibles au public sont indiqués), les
signatures microbiennes qu’ils reconnaissent, ainsi que les types de cellules
principales qui expriment ces récepteurs (la liste n’est pas exhaustive non
plus). |
Aucun
microorganisme ne possède toutes ces signatures en même temps. Certaines
bactéries ont une membrane externe riche en lipopolysaccharides (bactéries à
Gram-négatif)[70],
d’autres en sont dépourvues et affichent un peptidoglycane décoré d’acides
lipoteichoïques[71];
il y a des bactéries à Gram-négatif disposant d’un ou plusieurs flagelles,
tandis que d’autres n’en ont pas[72].
Bref, tu comprends que les microorganismes présentent des combinaisons de signatures microbiennes et que l’ensemble
diversifié de récepteurs innés permet de les détecter malgré cette diversité de
caractéristiques. Il en est de même des champignons, des parasites ou, surtout,
des virus (qui sont encore plus divers que les autres). Un même microorganisme
peut donc être “senti” par différents récepteurs en même temps. Par exemple,
une bactérie à Gram-négatif possédant un flagelle peut être perçue par les
récepteurs TLR4 (détection du LPS), par les récepteurs TLR5 (détection de la
flagelline) et, comme elle dispose aussi d’un peptidoglycane, par les
récepteurs NOD et TLR2-TLR6 (Fig. 6). Le récepteur TLR9 peut aussi détecter l’ADN
bactérien des cellules bactériennes mourantes. Il existe en plus d’autres
récepteurs intracellulaires du LPS, différents du TLR4, notamment chez les
macrophages qui phagocytent (avalent) les bactéries[73].
Certains
TLR et récepteurs NOD peuvent aussi détecter des signatures de microorganismes
très différents entre eux. Par exemple, le TLR4 qui reconnaît le LPS des
bactéries à Gram-négatif est aussi impliqué dans la détection des O-mannanes (des sucres complexes
constitués de mannose) décorant les protéines de surface des champignons. Le
récepteur NOD-2, qui s’associe à certains composants du peptidoglycane
bactérien, peut aussi reconnaître la chitine, sucre complexe de la paroi des
champignons[74].
En
somme, la diversité des récepteurs innés n’est pas inutilement redondante, mais
permet la reconnaissance d’une grande diversité de microorganismes en dépit du
fait que ces récepteurs ne perçoivent qu’un nombre limité d’éléments conservés.
Le nombre limité de récepteurs est, en quelque sorte, compensé par la
possibilité de percevoir une grande variété de combinaisons de signatures et
donc de microorganismes.
Processus permanent de reconnaissance
La
reconnaissance microbienne par les récepteurs de l’immunité innée couvre donc
un spectre très large de caractéristiques — des caractéristiques présentes, en
différentes combinaisons, chez tous
les micro-organismes, qu’ils soient
pathogènes ou non. Il n’y a, comme je l’ai déjà mentionné, pas de
différence entre ces deux catégories sémantiques en termes de signatures
moléculaires. Les bactéries à Gram-négatif, pathogènes ou non, possèdent toutes
du LPS, parfois un ou plusieurs flagelles, et sont détectées de la même
manière. Il en est de même des bactéries « à Gram-positif »,
pathogènes ou non, avec leurs signatures constituées de peptidoglycane et
d’acides lipoteichoïques. Les champignons non-pathogènes, de même, possèdent
aussi les b-glucanes, la chitine ou
les a-mannanes typiques des
parois fongiques (des polysaccharides) et qui sont détectés par les mêmes récepteurs que ceux
impliqués dans la reconnaissance de leurs homologues pathogènes.
Les
trillions de bactéries présentes dans nos intestins, membres permanents ou
transitoires du microbiote, possèdent ces mêmes signatures bactériennes, qui
sont bien évidemment perçues par les mêmes récepteurs — les TLRs, les NODs, etc
— par les cellules épithéliales intestinales, par les cellules dendritiques,
etc[75][76].
Les champignons du mycobiote, eux
aussi, sont ressentis en permanence par les mêmes récepteurs innés que ceux
intervenant dans la détection des versions pathogènes[77][78].
Ces microorganismes sont présents en
permanence et leur détection provoque bien évidemment le déclenchement des
processus immunitaires innés classiques (inflammation), mais aussi adaptatifs,
avec production d’anticorps spécifiques (on le verra plus tard).
C’est
donc bel et bien ce qui se produit en permanence dans nos intestins avec le
microbiote intestinal, mais aussi dans toutes les zones d’interaction avec nos
flores microbiennes (peau, muqueuse respiratoire, etc). Aucuns risques, donc, que le système immunitaire
soit en “pause” du point de vue du système immunitaire inné. Et nous
verrons un de ces jours que c’est d’autant plus vrai pour le système
immunitaire adaptatif.
Le
pathogène et le non-pathogène, une question de point de vue
En
fait, si un organisme pouvant entrer et survivre dans nos corps est
non-pathogène, c’est précisément parce
qu’il est reconnu et jugulé par
le système immunitaire. Il déclenche les mêmes mécanismes que dans le cas d’un
organisme pathogène. En général, l’immunité innée suffit à empêcher une
prolifération incontrôlée, ce qui nous conduit à qualifier un tel
microorganisme de non-pathogène. Le même raisonnement est valable pour le
système immunitaire adaptatif (qu’on verra plus tard). Les symptômes
pathologiques (de maladie) n’apparaissent pas car les mécanismes immunitaires
sont assez rapides et efficaces pour éviter l’activation d’une inflammation à
plus grande échelle. La prolifération microbienne et la dissémination du
microorganisme sont empêchées.
Note
ici que la réponse innée (mais aussi adaptative) est permanente vis-à-vis des microorganismes du microbiote.
L’inflammation permanente — qu’on devrait appeler “stimulation immunitaire
permanente” — causée de ces microorganismes est pourtant bien là, les réponses
immunitaires sont bel et bien enclenchées, mais tu ne perçois rien. Tu verras bientôt dans un futur billet qu’il y a une
différence fondamentale entre le fait de déclencher l’inflammation (les
mécanismes immunitaires) et déclencher des symptômes de maladie.
Les
microorganismes qu’on qualifie de pathogènes
(parce qu’ils déclenchent des symptômes de maladie) sont, en réalité, les microorganismes qui échappent
(temporairement) à ces mécanismes de réponse et qui induisent une réponse de
plus grande ampleur parce qu’ils prolifèrent — réponse de plus grande
échelle qui déclenche des symptômes physiologiques (fièvre, fatigue,
courbatures, etc.). De manière générale, les microorganismes pathogènes
possèdent les mêmes caractéristiques moléculaires que les autres, mais ils
disposent de moyens de masquer ces signatures moléculaires suffisamment
longtemps — ou un peu plus efficacement, sans l’empêcher totalement — pour
échapper temporairement à la détection innée, voire à la reconnaissance
spécifique par l’immunité adaptative. Ces capacités de masquage existent aussi
chez les non-pathogènes, mais ne les rend pas délétère pour autant. C’est la
combinaison de ces traits chez certains microorganismes qui les rend capables
de se multiplier en dépit des conditions défavorables induites par nos systèmes
physiologiques internes et/ou par la détection et la réponse immunitaires.
Certains microorganismes, par exemple, peuvent s’installer dans nos cellules et éviter la réponse immunitaire extracellulaire,
mais aussi esquiver en partie celle qui se produit dans nos cellules.
Les
virus pathogènes, par exemple, sont ceux qui peuvent 1) entrer dans nos
cellules, 2) s’y multiplier, 3) échapper à la réponse immunitaire assez
longtemps pour que leur multiplication et les conséquences de cette dernière
deviennent délétères. Il existe pléthore de virus non-pathogènes pour les
humains alors qu’ils possèdent toute la machinerie les rendant capables de se
multiplier au sein de nos cellules. Ils sont non-pathogènes du simple fait
qu’ils ne peuvent y entrer, rendant leur machinerie inutile dans ce contexte.
De la même manière, certains virus peuvent pénétrer nos cellules, mais ne s’y
multiplient pas, car les éléments qui permettraient leur cycle de réplication
n’y existent pas ou n’y sont pas exprimés d’une manière favorable ou encore
parce que l’immunité innée (dit antivirale dans ce contexte) est suffisamment
efficace pour que cela ne survienne pas. Car les récepteurs de l’immunité innée
peuvent aussi les détecter, même s’ils ne sont pas pathogènes : ils
possèdent les mêmes caractéristiques anormales que les autres virus semblables
mais pathogènes. Certaines bactéries peuvent également entrer dans nos cellules
et s’y dissimuler, les protégeant dans une certaine mesure de la réponse
immunitaire[79].
Ce qui est pathogène c’est ce qui
échappe à nos défenses. Par définition. Sinon, ils ne sont pas pathogènes. Par définition.
Le
microbiote est complexe. Chez les humains, des milliers d’espèces bactériennes
coexistent[80]
et on estime que le corps humain compte au moins autant de cellules
bactériennes qu’il compte de cellules “humaines”[81]. Mais en plus de cette
flore bactérienne, il faut compter le mycobiome[82][83],
l’ensemble des champignons transitoires ou résidents, l’archéome[84],
l’ensemble des archées [prononcer arkées]
transitoires ou permanentes (des microorganismes qui ne sont ni des bactéries,
ni des eucaryotes) et le virome[85][86],
l’ensemble des virus transitoires ou permanents. On parle ici de
microorganismes présents chez des individus sains, asymptomatiques, et donc de
ce qu’on appelle la flore microbienne normale.
Rien
que sur la peau, on trouve des milliers d’espèces bactériennes, fongiques
(champignons) et virales[87],
composition qui change beaucoup selon les endroits qu’on fréquente, l’hygiène
corporelle et sa fréquence, les activités de la journée, la zone dermique
considérée, l’humidité de la zone corporelle, la fréquence et la durée des
contacts avec des surfaces, des animaux, des plantes, d’autres humains, etc[88][89].
La cavité buccale[90],
le système respiratoire[91]
et d’autres cavités ouvertes directement sur l’extérieur[92][93]
sont elles aussi colonisées par une microflore absolument gigantesque, elle
aussi dynamique et changeante (alimentation, qualité de l’air, etc.). Il n’en
est pas moins que certaines espèces des voies respiratoires sont des résidents
permanents, comme certains Streptococcus,
des Corynebacterium et autres Staphylococcus aureus (staphylocoque
doré)[94].
Le microbiote intestinal est un peu plus stable, mais extrêmement diversifié[95][96]:
bactéries, archées, protozoaires, champignons et virus. L’intestin est aussi — en plus d’un organe d’assimilation
des nutriments — un organe de “stockage” des microbes.
Il
existe d’étonnants virus du microbiote humain, les Anelloviridae, capable d’entrer dans certaines cellules (dont on
ignore l’identité), de s’y multiplier et de se disséminer dans l’organisme (on
les détecte dans de nombreux organes, dans la circulation sanguine, etc.[97]),
et qui infectent pratiquement toute la population humaine mondiale[98].
Leur présence ne semble pas causer de symptômes notables, ni de maladies, en
dépit du fait qu’ils sont détectés par l’immunité innée antivirale. Ils sont
détectés, certes, mais disposent de mécanismes d’échappement immunitaire assez
efficaces[99]
leur permettant de persister toute la vie chez tous les êtres humains. Leur
taux sanguin augmente beaucoup chez les personnes immunodéprimées, indiquant tout
de même une certaine forme de contrôle
immunitaire. Ce genre de virus existe aussi chez beaucoup d’autres mammifères.
Bien qu’aucune maladie n’ait été aujourd’hui associée à ces virus, et qu’ils
participent probablement à la modulation de l’immunité, ils pourraient avoir
des effets en situation d’immunosuppression ou de dysfonction immunitaire, de
fatigue chronique, et on sait que leur abondance change pendant une infection
par le SARS-CoV-2 et dans les cas de Covid-long[100][101][102][103],
à tel point qu’ils sont de bons marqueurs de gravité.
Même si beaucoup de bactéries sont des résidentes
permanentes et confinées par les barrières des muqueuses faisant d’eux des
commensaux, celles qui colonisent les voies respiratoires deviennent pathogènes
lorsque ces voies sont altérées par les conséquences d’une infection
respiratoire — pas nécessairement grave — comme après une infection virale
respiratoire[104][105][106]. Même s’ils étaient
présents depuis toujours, ils peuvent provoquer de graves infections invasives
car, une fois la barrière des muqueuses passée, ils disposent de mécanismes
d’échappement immunitaire et de facteurs de virulence qui les rendent dangereux
une fois entrés. La reconnaissance immunitaire avait bien lieu lorsqu’ils se trouvaient
dans les voies respiratoires, car ils possèdent des signatures typiques de
celles détectées par les récepteurs de l’immunité innée, mais cela ne confère
guère d’avantages une fois qu’ils ont passé la barrière des muqueuses, car leur
capacité d’invasion est nettement supérieure à celle d’élimination par le
système immunitaire.
De la même manière, lorsque le système immunitaire
est altéré, certains membres du microbiote peuvent devenir pathogènes, car en
l’absence de limitations imposées par lui, ils prolifèrent de façon incontrôlée[107].
Chez les personnes infectées par le virus de l’immunodéficience humaine (VIH),
lorsque les cellules immunitaires deviennent trop peu nombreuses à cause de
l’infection virale qui les détruit progressivement, des microorganismes
normalement résidents — disons normalement neutres
— deviennent néfastes, pathogènes, et causent des maladies dites opportunistes:
le champignon Candida albicans,
normalement présent sur la peau, les muqueuses, dans le tube digestif devient
ainsi hors de contrôle[108].
Les mêmes problèmes surviennent avec d’autres maladies qui causent des
dysfonctions immunitaires. La menace provient alors de microorganismes qui
étaient jusque là résidents, mais aussi de microorganismes extérieurs qui
apparaissaient comme “non-pathogènes” jusqu’à ce que les dysfonctions
immunitaires surviennent[109].
Bref, le
caractère pathogène ou non-pathogène dépend à la fois des caractéristiques ou
des combinaisons de caractéristiques du microorganisme qui interagit avec nous,
mais aussi des conditions internes et immunitaires de l’hôte. La réponse
immunitaire est donc elle-même un critère, puisque, comme je l’ai déjà
mentionné, une diminution des capacités immunitaires (maladie immunitaire,
immunosuppression, immunodépression, dysfonction immunitaire, présence d’autres
microorganismes, carences, etc.) peut rendre des microorganismes normalement
résidents et/ou inoffensifs, dangereux. La pathogénie est une propriété
contextuelle, à la fois pour le microorganisme et pour l’hôte qu’il visite et est une question d’interaction.
Détection des
dommages et de cellules anormales
Le principe de reconnaissance des
cellules endommagées, anormales (cancéreuses, mortes, etc.) repose sur les
mêmes mécanismes. Il existe des récepteurs reconnaissant des caractéristiques anormales
(Damage-Associated
Molecular Patterns, DAMPs) et équipant les cellules
immunitaires, lesquelles répondent également en déclenchant l’inflammation. Ces
récepteurs sont en grande partie les mêmes que ceux qui permettent de détecter
les microorganismes[110].
Les MAMPs et les DAMPs sont des stimuli, des éléments qui activent (stimulent) les cellules immunitaires
spécialisées ou les cellules qui possèdent les détecteurs appropriés.
L’inflammation est la marque de cette activation[111].
Cette dernière peut être sensible (comme lorsqu’on est malade, que le système
est déclenché à plein, à grande échelle), ou indolore, lorsque les choses se
passent sur une très courte échelle de temps et très localement. Le microbiote,
les microorganismes de passage, les cellules vieillissantes, mortes,
endommagées, les molécules diverses et variées qui passent par là, toutes ces entités reconnaissables par
les cellules exprimant des récepteurs innés dédiés sont des stimuli. Pense, par
exemple, que les allergies sont des
stimulations dues à des molécules, parfois bien plus petites que des
microorganismes, qui témoignent de ce genre de reconnaissance — même si dans ce
cas, il s’agit d’une sur-activation du système immunitaire. Car oui, ce que
nous mangeons est aussi détecté par le système immunitaire, éléments qui,
plusieurs fois par jour “stimulent” donc la machinerie.
En conclusion
Le système immunitaire
est sollicité en permanence. Toute
détection d’une caractéristique anormale déclenche l’inflammation (la mise en
route d’une réponse immunitaire) mais cela ne se propage pas toujours à la même
échelle. Seul un déclenchement à grande échelle provoque les fameux symptômes de
maladie.
REFERENCES
[1] Je fais évidemment ici référence
aux « théories » qui parlent de « dette immunitaire » et
qui prétendent que le système immunitaire serait « sous-stimulé »
lorsqu’on n’est pas exposé aux pathogènes et donc aux maladies.
[2] Cassa Macedo A, Oliveira Vilela de
Faria A, Ghezzi P. Boosting the Immune System, From Science to Myth: Analysis
the Infosphere With Google. Front Med (Lausanne). 2019 Jul 25;6:165. doi:
10.3389/fmed.2019.00165.
[3]
Locey KJ, Lennon JT. Scaling laws predict global microbial diversity. Proc Natl
Acad Sci U S A. 2016 May 24;113(21):5970-5. doi: 10.1073/pnas.1521291113.
[4] Levin BR, Antia R. Why we don't get
sick: the within-host population dynamics of bacterial infections. Science.
2001 May 11;292(5519):1112-5. doi: 10.1126/science.1058879
[5]
Martínez JL. Bacterial pathogens: from natural ecosystems to human hosts. Environ
Microbiol. 2013
Feb;15(2):325-33. doi: 10.1111/j.1462-2920.2012.02837.x.
[6] Rohmer L, Hocquet D, Miller SI. Are
pathogenic bacteria just looking for food? Metabolism and microbial
pathogenesis. Trends Microbiol. 2011
Jul;19(7):341-8. doi: 10.1016/j.tim.2011.04.003.
[7] Fischer, F. and De
Reuse, H. (2016) Adaptation
of Helicobacter pylori metabolism to persistent
gastric colonization in “Helicobacter pylori Research: From
Bench to Bedside”, Editors: Yamaoka,
Y. and Backert,
S (eds). Publisher:
Springer.
[8] Walter EJ, Hanna-Jumma S,
Carraretto M, Forni L. The pathophysiological basis and consequences of fever. Crit
Care. 2016 Jul 14;20(1):200. doi: 10.1186/s13054-016-1375-5.
[9] Belon L, Skidmore P, Mehra R,
Walter E. Effect of a fever in viral infections - the 'Goldilocks' phenomenon?
World J Clin Cases. 2021 Jan 16;9(2):296-307. doi: 10.12998/wjcc.v9.i2.296.
[10] Casadevall A. Thermal Restriction
as an Antimicrobial Function of Fever. PLoS Pathog. 2016 May 5;12(5):e1005577.
doi: 10.1371/journal.ppat.1005577.
[11] Evans SS, Repasky EA, Fisher DT.
Fever and the thermal regulation of immunity: the immune system feels the heat.
Nat Rev Immunol. 2015 Jun;15(6):335-49. doi: 10.1038/nri3843.
[12] Zygiel EM, Nolan EM. Transition Metal Sequestration by the Host-Defense Protein Calprotectin.
Annu Rev Biochem. 2018 Jun 20;87:621-643. doi:
10.1146/annurev-biochem-062917-012312.
[13] M. I. Hood, E. P. Skaar (2012) Nutritional immunity:
transition metals at the pathogen–host interface. Nat. Rev Microbiol 10,
525–537.
[14] Murdoch CC, Skaar EP.
Nutritional immunity: the battle for nutrient metals at the host-pathogen
interface. Nat Rev Microbiol. 2022 May 31:1–14. doi: 10.1038/s41579-022-00745-6.
[15]
Nicholson LB. Op. Cit.
[16]
Yuan S, Tao X, Huang S, Chen S, Xu A. Comparative immune systems in animals. Annu Rev Anim Biosci. 2014;2:235-58.
doi: 10.1146/annurev-animal-031412-103634.
[17]
Spoel SH, Dong X. How do plants achieve immunity? Defence without specialized
immune cells. Nat Rev Immunol. 2012;12(2):89-100.
doi: 10.1038/nri3141.
[18]
Gaspar ML, Pawlowska TE. Innate immunity in fungi: Is regulated cell death
involved? PLoS Pathog. 2022;18(5):e1010460. doi:
10.1371/journal.ppat.1010460.
[19] Marraffini LA. CRISPR-Cas
immunity in prokaryotes. Nature.
2015;526(7571):55-61. doi: 10.1038/nature15386.
[20]
Hirano M, Das S, Guo P, Cooper MD. The evolution of adaptive immunity in
vertebrates. Adv Immunol. 2011; 109:125-57.
doi: 10.1016/B978-0-12-387664-5.00004-2
[21]
Sirisinha S. Evolutionary insights into the origin of innate and adaptive
immune systems: different shades of grey. Asian
Pac J Allergy Immunol. 2014; 32(1):3-15.
[22]
Flajnik MF, Kasahara M. Op. cit.
[23]
Kaufman J. Unfinished Business: Evolution of the MHC and the Adaptive Immune
System of Jawed Vertebrates. Annu Rev Immunol. 2018 Apr 26;36:383-409. doi:
10.1146/annurev-immunol-051116-052450.
[24]
Chaplin DD. Overview of the immune response. J Allergy Clin Immunol. 2010
Feb;125(2 Suppl 2):S3-23. doi: 10.1016/j.jaci.2009.12.980.
[25] Gasteiger G, D'Osualdo A, Schubert
DA, Weber A, Bruscia EM, Hartl D. Cellular Innate Immunity: An Old Game with
New Players. J Innate Immun. 2017;9(2):111-125. doi: 10.1159/000453397.
[26]
Nagasawa M, Spits H, Ros XR. Innate
Lymphoid Cells (ILCs): Cytokine Hubs Regulating Immunity and Tissue
Homeostasis. Cold Spring Harb Perspect Biol. 2018 Dec 3;10(12):a030304. doi:
10.1101/cshperspect.a030304.
[27]
Crinier A, Viant C, Girard-Madoux M, Vivier É. Les cellules lymphoïdes innées
[Innate lymphoid cells]. Med
Sci (Paris). 2017 May;33(5):534-542. French. doi: 10.1051/medsci/20173305018.
[28] Sender R, Weiss Y, Navon Y, Milo I,
Azulay N, Keren L, Fuchs S, Ben-Zvi D, Noor E, Milo R. The total mass, number,
and distribution of immune cells in the human body. Proc Natl Acad Sci U S A.
2023 Oct 31;120(44):e2308511120. doi: 10.1073/pnas.2308511120.
[29] Hatton IA, Galbraith ED, Merleau
NSC, Miettinen TP, Smith BM, Shander JA. The human cell count and size
distribution. Proc Natl Acad Sci U S A. 2023 Sep 26;120(39):e2303077120. doi:
10.1073/pnas.2303077120.
[30] Powell N, Walker MM, Talley NJ. The
mucosal immune system: master regulator of bidirectional gut-brain communications.
Nat Rev Gastroenterol Hepatol. 2017 Mar;14(3):143-159. doi:
10.1038/nrgastro.2016.191.
[31] Chu H, Mazmanian SK. Innate
immune recognition of the microbiota promotes host-microbial symbiosis. Nat Immunol. 2013;14(7):668-75. doi:
10.1038/ni.2635.
[32] Méthot PO, Alizon S. What is
a pathogen? Toward a process view of host-parasite interactions. Virulence. 2014;5(8):775-85. doi:
10.4161/21505594.2014.960726.
[33] Maehle AH. A binding question: the
evolution of the receptor concept. Endeavour. 2009 Dec;33(4):135-40. doi:
10.1016/j.endeavour.2009.09.001.
[34] Silverstein,
Arthur M. A History
of Immunology, Second Edition (2009, Academic Press), 2009.
[35] Jarmoskaite I, AlSadhan I,
Vaidyanathan PP, Herschlag D. How to measure and evaluate binding affinities. Elife.
2020 Aug 6;9:e57264. doi: 10.7554/eLife.57264.
[36]
Rubartelli A, Lotze MT. Inside, outside, upside down: damage-associated
molecular-pattern molecules (DAMPs) and redox. Trends Immunol. 2007
Oct;28(10):429-36. doi: 10.1016/j.it.2007.08.004.
[37]
Tang D, Kang R, Coyne CB, Zeh HJ, Lotze MT. PAMPs and DAMPs: signal 0s that
spur autophagy and immunity. Immunol Rev. 2012 Sep;249(1):158-75. doi:
10.1111/j.1600-065X.2012.01146.x.
[38]
Hatt H. Molecular and cellular basis of human olfaction. Chem Biodivers. 2004
Dec;1(12):1857-69. doi: 10.1002/cbdv.200490142.
[39]
Gravina SA, Yep GL, Khan M. Human biology of taste. Ann Saudi Med. 2013
May-Jun;33(3):217-22. doi: 10.5144/0256-4947.2013.217.
[40]
Yoshioka T, Sakakibara M. Physical aspects of sensory transduction on seeing,
hearing and smelling. Biophysics (Nagoya-shi). 2013;9:183-91.
[41]
Roy S, Nagrale P. Encoding the Photoreceptors of the Human Eye. Cureus. 2022
Oct 10;14(10):e30125. doi: 10.7759/cureus.30125.
[42]
Xiao R, Xu XZS. Temperature Sensation: From Molecular Thermosensors to Neural
Circuits and Coding Principles. Annu Rev Physiol. 2021 Feb 10;83:205-230. doi:
10.1146/annurev-physiol-031220-095215.
[43]
Roudaut Y, Lonigro A, Coste B, Hao J, Delmas P, Crest M. Touch sense:
functional organization and molecular determinants of mechanosensitive
receptors. Channels (Austin). 2012 Jul-Aug;6(4):234-45.
[44]
Dubin AE, Patapoutian A. Nociceptors: the sensors of the pain pathway. J Clin
Invest. 2010 Nov;120(11):3760-72. doi: 10.1172/JCI42843.
[45]
L Kiss A. Inflammation in Focus: The Beginning and the End. Pathol Oncol Res.
2022 Jan 4;27:1610136. doi: 10.3389/pore.2021.1610136.
[46] Silverstein, 2009, Op. Cit.
[47]
Annunziato F, Romagnani C, Romagnani S. The 3 major types of innate and
adaptive cell-mediated effector immunity. J Allergy Clin Immunol. 2015
Mar;135(3):626-35. doi: 10.1016/j.jaci.2014.11.001.
[48] Li D, Wu M. Pattern recognition
receptors in health and diseases. Signal Transduct Target Ther. 2021 Aug
4;6(1):291. doi: 10.1038/s41392-021-00687-0.
[49]
Anthoney N, Foldi I, Hidalgo A. Toll and Toll-like receptor signalling in
development. Development. 2018 Apr 25;145(9):dev156018. doi:
10.1242/dev.156018.
[50]
Buckley KM, Yoder JA. The evolution of innate immune receptors: investigating
the diversity, distribution, and phylogeny of immune recognition across
eukaryotes. Immunogenetics. 2022 Feb;74(1):1-4. doi:
10.1007/s00251-021-01243-4.
[51]
Kanneganti TD. Intracellular innate immune receptors: Life inside the cell.
Immunol Rev. 2020 Sep;297(1):5-12. doi: 10.1111/imr.12912.
[52]
Liston A, Humblet-Baron S, Duffy D, Goris A. Human immune diversity: from
evolution to modernity. Nat Immunol. 2021 Dec;22(12):1479-1489. doi: 10.1038/s41590-021-01058-1.
[53]
Vilches C, Parham P. KIR: diverse, rapidly evolving receptors of innate and
adaptive immunity. Annu Rev Immunol. 2002;20:217-51. doi:
10.1146/annurev.immunol.20.092501.134942.
[54]
Dębińska A, Danielewicz H, Drabik-Chamerska A, Kalita D, Boznański A. Genetic
polymorphisms in pattern recognition receptors are associated with allergic
diseases through gene-gene interactions. Adv Clin Exp Med. 2019
Aug;28(8):1087-1094. doi: 10.17219/acem/104538.
[55]
Garde S, Chodisetti PK, Reddy M. Peptidoglycan: Structure, Synthesis, and
Regulation. EcoSal Plus. 2021 Jan;9(2). doi: 10.1128/ecosalplus.ESP-0010-2020.
[56]
Chou WC, Jha S, Linhoff MW, Ting JP. The NLR gene family: from discovery to
present day. Nat Rev Immunol. 2023 Oct;23(10):635-654. doi:
10.1038/s41577-023-00849-x.
[57]
Babamale AO, Chen ST. Nod-like Receptors: Critical Intracellular Sensors for
Host Protection and Cell Death in Microbial and Parasitic Infections. Int J Mol
Sci. 2021 Oct 22;22(21):11398. doi: 10.3390/ijms222111398.
[58]
Motta V, Soares F, Sun T, Philpott DJ. NOD-like receptors: versatile cytosolic
sentinels. Physiol Rev. 2015 Jan;95(1):149-78. doi: 10.1152/physrev.00009.2014.
[59]
Molinaro A, Holst O, Di Lorenzo F, Callaghan M, Nurisso A, D'Errico G,
Zamyatina A, Peri F, Berisio R, Jerala R, Jiménez-Barbero J, Silipo A,
Martín-Santamaría S. Chemistry of lipid A: at the heart of innate immunity.
Chemistry. 2015 Jan 7;21(2):500-19. doi: 10.1002/chem.201403923.
[60]
Di Lorenzo F, Duda KA, Lanzetta R, Silipo A, De Castro C, Molinaro A. A Journey
from Structure to Function of Bacterial Lipopolysaccharides. Chem Rev. 2022 Oct
26;122(20):15767-15821. doi: 10.1021/acs.chemrev.0c01321.
[61]
Park BS, Song DH, Kim HM, Choi BS, Lee H, Lee JO. The structural basis of
lipopolysaccharide recognition by the TLR4-MD-2 complex. Nature. 2009 Apr
30;458(7242):1191-5. doi: 10.1038/nature07830.
[62]
Sameer AS, Nissar S. Toll-Like Receptors (TLRs): Structure, Functions,
Signaling, and Role of Their Polymorphisms in Colorectal Cancer Susceptibility.
Biomed Res Int. 2021 Sep 12;2021:1157023. doi: 10.1155/2021/1157023.
[63] Kato T, Makino F, Miyata T, Horváth
P, Namba K. Structure of the native supercoiled flagellar hook as a universal
joint. Nat Commun. 2019 Nov 22;10(1):5295. doi: 10.1038/s41467-019-13252-9.
[64] Hajam IA, Dar PA, Shahnawaz I,
Jaume JC, Lee JH. Bacterial flagellin-a potent immunomodulatory agent. Exp Mol
Med. 2017 Sep 1;49(9):e373. doi: 10.1038/emm.2017.172.
[65]
Patin EC, Thompson A, Orr SJ. Pattern recognition receptors in fungal immunity.
Semin Cell Dev Biol. 2019 May;89:24-33. doi: 10.1016/j.semcdb.2018.03.003.
[66] Gow NAR, Latge JP, Munro CA. The
Fungal Cell Wall: Structure, Biosynthesis, and Function. Microbiol Spectr. 2017
May;5(3). doi: 10.1128/microbiolspec.FUNK-0035-2016.
[67] Nikolakopoulou C, Willment JA,
Brown GD. C-Type Lectin Receptors in Antifungal Immunity. Adv Exp Med Biol.
2020;1204:1-30. doi: 10.1007/978-981-15-1580-4_1.
[68]
Carty M, Guy C, Bowie AG. Detection of Viral Infections by Innate Immunity. Biochem Pharmacol. 2021
Jan;183:114316. doi: 10.1016/j.bcp.2020.114316.
[69] Tel que formulé par Charles
Jeneway en 1989: Janeway CA Jr. Approaching the asymptote?
Evolution and revolution in immunology. Cold Spring Harb Symp Quant Biol.
1989;54 Pt 1:1-13. doi: 10.1101/sqb.1989.054.01.003.
[70] Henderson JC, Zimmerman SM, Crofts
AA, Boll JM, Kuhns LG, Herrera CM, Trent MS. The Power of Asymmetry:
Architecture and Assembly of the Gram-Negative Outer Membrane Lipid Bilayer.
Annu Rev Microbiol. 2016 Sep 8;70:255-78. doi: 10.1146/annurev-micro-102215-095308.
[71] Percy MG, Gründling A. Lipoteichoic
acid synthesis and function in gram-positive bacteria. Annu Rev Microbiol.
2014;68:81-100. doi: 10.1146/annurev-micro-091213-112949.
[72] Chu J, Liu J, Hoover TR.
Phylogenetic Distribution, Ultrastructure, and Function of Bacterial Flagellar
Sheaths. Biomolecules. 2020 Feb 27;10(3):363. doi: 10.3390/biom10030363.
[73] Demon D, Vande Walle L, Lamkanfi M.
Sensing the enemy within: how macrophages detect intracellular Gram-negative
bacteria. Trends Biochem Sci. 2014 Dec;39(12):574-6. doi:
10.1016/j.tibs.2014.10.006.
[74] Netea MG, Joosten LA, van der Meer
JW, Kullberg BJ, van de Veerdonk FL. Immune defence against Candida fungal
infections. Nat Rev Immunol. 2015 Oct;15(10):630-42. doi: 10.1038/nri3897.
[75] Jordan CKI, Brown RL, Larkinson
MLY, Sequeira RP, Edwards AM, Clarke TB. Symbiotic Firmicutes establish
mutualism with the host via innate tolerance and resistance to control systemic
immunity. Cell Host Microbe. 2023 Sep 13;31(9):1433-1449.e9. doi:
10.1016/j.chom.2023.07.008.
[76] Guzman-Bautista ER, Suzuki K, Asami
S, Fagarasan S. Bacteria-immune cells dialog and the homeostasis of the
systems. Curr Opin Immunol. 2020 Oct;66:82-89. doi: 10.1016/j.coi.2020.05.010.
[77] Underhill DM, Iliev ID. The
mycobiota: interactions between commensal fungi and the host immune system. Nat
Rev Immunol. 2014 Jun;14(6):405-16. doi: 10.1038/nri3684.
[78] Limon JJ, Skalski JH, Underhill DM.
Commensal Fungi in Health and Disease. Cell Host Microbe. 2017 Aug
9;22(2):156-165. doi: 10.1016/j.chom.2017.07.002.
[79] Thakur A, Mikkelsen H, Jungersen G.
Intracellular Pathogens: Host Immunity and Microbial Persistence Strategies. J
Immunol Res. 2019 Apr 14;2019:1356540. doi: 10.1155/2019/1356540.
[80] Gilbert JA, Blaser MJ, Caporaso JG,
Jansson JK, Lynch SV, Knight R. Current understanding of the human microbiome.
Nat Med. 2018 Apr 10;24(4):392-400. doi: 10.1038/nm.4517.
[81] Sender R, Fuchs S, Milo R. Revised
Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol.
2016 Aug 19;14(8):e1002533. doi: 10.1371/journal.pbio.1002533.
[82] Bonfante P, Venice F, Lanfranco L.
The mycobiota: fungi take their place between plants and bacteria. Curr Opin
Microbiol. 2019 Jun;49:18-25. doi: 10.1016/j.mib.2019.08.004.
[83] Ost KS, Round JL. Commensal fungi
in intestinal health and disease. Nat Rev Gastroenterol Hepatol. 2023
Nov;20(11):723-734. doi: 10.1038/s41575-023-00816-w.
[84] Chibani CM, Mahnert A, Borrel G,
Almeida A, Werner A, Brugère JF, Gribaldo S, Finn RD, Schmitz RA,
Moissl-Eichinger C. A catalogue of 1,167 genomes from the human gut archaeome.
Nat Microbiol. 2022 Jan;7(1):48-61. doi: 10.1038/s41564-021-01020-9. Epub 2021
Dec 30. Erratum in: Nat Microbiol. 2022 Feb;7(2):339.
[85] Stockdale SR, Hill C. Progress and
prospects of the healthy human gut virome. Curr Opin Virol. 2021
Dec;51:164-171. doi: 10.1016/j.coviro.2021.10.001.
[86] Liang G, Bushman FD. The human
virome: assembly, composition and host interactions. Nat Rev Microbiol. 2021
Aug;19(8):514-527. doi: 10.1038/s41579-021-00536-5.
[87] Byrd AL, Belkaid Y, Segre JA. The
human skin microbiome. Nat Rev Microbiol. 2018 Mar;16(3):143-155. doi:
10.1038/nrmicro.2017.157.
[88] Saheb Kashaf S, Proctor DM, Deming
C, Saary P, Hölzer M; NISC Comparative Sequencing Program; Taylor ME, Kong HH,
Segre JA, Almeida A, Finn RD. Integrating cultivation and metagenomics for a
multi-kingdom view of skin microbiome diversity and functions. Nat Microbiol.
2022 Jan;7(1):169-179. doi: 10.1038/s41564-021-01011-w.
[89] Grice EA, Segre JA. The skin
microbiome. Nat Rev Microbiol. 2011 Apr;9(4):244-53. doi: 10.1038/nrmicro2537. Erratum
in: Nat Rev Microbiol. 2011 Aug;9(8):626.
[90] Willis JR, Gabaldón T. The Human
Oral Microbiome in Health and Disease: From Sequences to Ecosystems.
Microorganisms. 2020 Feb 23;8(2):308. doi: 10.3390/microorganisms8020308.
[91] Man WH, de Steenhuijsen Piters WA,
Bogaert D. The microbiota of the respiratory tract: gatekeeper to respiratory
health. Nat Rev Microbiol. 2017 May;15(5):259-270. doi:
10.1038/nrmicro.2017.14.
[92] Mendling W. Vaginal Microbiota. Adv
Exp Med Biol. 2016;902:83-93. doi: 10.1007/978-3-319-31248-4_6.
[93] Jones-Freeman B, Chonwerawong M,
Marcelino VR, Deshpande AV, Forster SC, Starkey MR. The microbiome and host
mucosal interactions in urinary tract diseases. Mucosal Immunol. 2021
Jul;14(4):779-792. doi: 10.1038/s41385-020-00372-5.
[94] Levy RM, Leyden JJ, Margolis DJ.
Colonisation rates of Streptococcus pyogenes and Staphylococcus aureus in the
oropharynx of a young adult population. Clin Microbiol Infect. 2005
Feb;11(2):153-5. doi: 10.1111/j.1469-0691.2004.01042.x.
[95] Rinninella E, Raoul P, Cintoni M,
Franceschi F, Miggiano GAD, Gasbarrini A, Mele MC. What is the Healthy Gut
Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and
Diseases. Microorganisms. 2019 Jan 10;7(1):14. doi:
10.3390/microorganisms7010014.
[96] Fan Y, Pedersen O. Gut microbiota
in human metabolic health and disease. Nat Rev Microbiol. 2021 Jan;19(1):55-71.
doi: 10.1038/s41579-020-0433-9.
[97] Pyöriä L, Pratas D, Toppinen M,
Hedman K, Sajantila A, Perdomo MF. Unmasking the tissue-resident eukaryotic DNA
virome in humans. Nucleic Acids Res. 2023 Apr 24;51(7):3223-3239. doi:
10.1093/nar/gkad199.
[98] Taylor LJ, Keeler EL, Bushman FD,
Collman RG. The enigmatic roles of Anelloviridae and Redondoviridae in humans.
Curr Opin Virol. 2022 Aug;55:101248. doi: 10.1016/j.coviro.2022.101248. Taylor
LJ, Keeler EL, Bushman FD, Collman RG. The enigmatic roles of Anelloviridae and
Redondoviridae in humans. Curr Opin Virol. 2022 Aug;55:101248. doi:
10.1016/j.coviro.2022.101248.
[99] Liou SH, Boggavarapu R, Cohen NR,
Zhang Y, Sharma I, Zeheb L, Mukund Acharekar N, Rodgers HD, Islam S, Pitts J,
Arze C, Swaminathan H, Yozwiak N, Ong T, Hajjar RJ, Chang Y, Swanson KA,
Delagrave S. Structure of anellovirus-like particles reveal a mechanism for
immune evasion. Nat Commun. 2024 Aug 22;15(1):7219. doi:
10.1038/s41467-024-51064-8.
[100] Sabbaghian M, Gheitasi H,
Shekarchi AA, Tavakoli A, Poortahmasebi V. The mysterious anelloviruses:
investigating its role in human diseases. BMC Microbiol. 2024 Jan 29;24(1):40.
doi: 10.1186/s12866-024-03187-7.
[101] Buonsenso D, Tantisira KG. Long
COVID and SARS-CoV-2 persistence: new answers, more questions. Lancet Infect
Dis. 2024 Aug;24(8):796-798. doi: 10.1016/S1473-3099(24)00216-0.
[102] Sasa N, Kojima S, Koide R, Hasegawa
T, Namkoong H, Hirota T, Watanabe R, Nakamura Y, Oguro-Igashira E, Ogawa K,
Yata T, Sonehara K, Yamamoto K, Kishikawa T, Sakaue S, Edahiro R, Shirai Y,
Maeda Y, Nii T, Chubachi S, Tanaka H, Yabukami H, Suzuki A, Nakajima K, Arase
N, Okamoto T, Nishikawa R, Namba S, Naito T, Miyagawa I, Tanaka H, Ueno M,
Ishitsuka Y, Furuta J, Kunimoto K, Kajihara I, Fukushima S, Miyachi H, Matsue
H, Kamata M, Momose M, Bito T, Nagai H, Ikeda T, Horikawa T, Adachi A,
Matsubara T, Ikumi K, Nishida E, Nakagawa I, Yagita-Sakamaki M, Yoshimura M,
Ohshima S, Kinoshita M, Ito S, Arai T, Hirose M, Tanino Y, Nikaido T, Ichiwata
T, Ohkouchi S, Hirano T, Takada T, Tazawa R, Morimoto K, Takaki M, Konno S,
Suzuki M, Tomii K, Nakagawa A, Handa T, Tanizawa K, Ishii H, Ishida M, Kato T,
Takeda N, Yokomura K, Matsui T, Uchida A, Inoue H, Imaizumi K, Goto Y, Kida H,
Fujisawa T, Suda T, Yamada T, Satake Y, Ibata H, Saigusa M, Shirai T, Hizawa N,
Nakata K; Japan COVID-19 Task Force; Imafuku S, Tada Y, Asano Y, Sato S,
Nishigori C, Jinnin M, Ihn H, Asahina A, Saeki H, Kawamura T, Shimada S,
Katayama I, Poisner HM, Mack TM, Bick AG, Higasa K, Okuno T, Mochizuki H, Ishii
M, Koike R, Kimura A, Noguchi E, Sano S, Inohara H, Fujimoto M, Inoue Y,
Yamaguchi E, Ogawa S, Kanai T, Morita A, Matsuda F, Tamari M, Kumanogoh A,
Tanaka Y, Ohmura K, Fukunaga K, Imoto S, Miyano S, Parrish NF, Okada Y. Blood
DNA virome associates with autoimmune diseases and COVID-19. Nat Genet. 2025
Jan;57(1):65-79. doi: 10.1038/s41588-024-02022-z.
[103] Maguire C, Chen J, Rouphael N,
Pickering H, Phan HV, Glascock A, Chu V, Dandekar R, Corry D, Kheradmand F,
Baden LR, Selaky R, McComsey GA, Haddad EK, Cairns CB, Pulendran B,
Fernandez-Sesma A, Simon V, Metcalf JP, Higuita NIA, Messer WB, David MM,
Nadeau KC, Kraft M, Bime C, Schaenman J, Erle D, Calfee CS, Atkinson MA,
Brackenridge SC, Ehrlich LIR, Montgomery RR, Shaw AC, Hough CL, Geng LN, Hafler
DA, Augustine AD, Becker PM, Peters B, Ozonoff A, Kim-Schulze SH, Krammer F,
Bosinger S, Eckalbar W, Altman MC, Wilson M, Guan L, Kleinstein SH; IMPACC
Network; Smolen KK, Reed EF, Levy O, Maecker H, Hunt P, Steen H, Diray-Arce J,
Langelier CR, Melamed E. Chronic Viral Reactivation and Associated Host Immune
Response and Clinical Outcomes in Acute COVID-19 and Post-Acute Sequelae of
COVID-19. bioRxiv [Preprint]. 2024 Nov 16:2024.11.14.622799. doi:
10.1101/2024.11.14.622799.
[104] Hament JM, Kimpen JL, Fleer A,
Wolfs TF. Respiratory viral infection predisposing for bacterial disease: a
concise review. FEMS Immunol Med Microbiol. 1999 Dec;26(3-4):189-95. doi:
10.1111/j.1574-695X.1999.tb01389.x.
[105] Hanada S, Pirzadeh M, Carver KY,
Deng JC. Respiratory Viral Infection-Induced Microbiome Alterations and
Secondary Bacterial Pneumonia. Front Immunol. 2018 Nov 16;9:2640. doi:
10.3389/fimmu.2018.02640.
[106]
Rossi GA, Fanous H, Colin AA. Viral
strategies predisposing to respiratory bacterial superinfections. Pediatr
Pulmonol. 2020 Apr;55(4):1061-1073. doi: 10.1002/ppul.24699.
[107]
Zheng D, Liwinski T, Elinav E. Interaction between microbiota and immunity in
health and disease. Cell Res.
2020;30(6):492-506. doi: 10.1038/s41422-020-0332-7.
[108]
Cassone A, Cauda R. Candida and candidiasis in HIV-infected patients: where
commensalism, opportunistic behavior and frank pathogenicity lose their
borders. AIDS. 2012;26(12):1457-72.
doi: 10.1097/QAD.0b013e3283536ba8.
[109]
Dropulic LK, Lederman HM. Overview of Infections in the Immunocompromised Host.
Microbiol Spectr.
2016;4(4):10.1128/microbiolspec.DMIH2-0026-2016. doi:
10.1128/microbiolspec.DMIH2-0026-2016.
[110] Rubartelli A, Lotze MT.
Inside, outside, upside down: damage-associated molecular-pattern molecules
(DAMPs) and redox. Trends Immunol. 2007 Oct;28(10):429-36. doi:
10.1016/j.it.2007.08.004.
[111]
Akira S, Uematsu S, Takeuchi O. Pathogen recognition and innate immunity. Cell.
2006 Feb 24;124(4):783-801. doi: 10.1016/j.cell.2006.02.015